Variación morfológica de la mandíbula en poblaciones del Pleistoceno terminal al Holoceno tardío en México
Rocío Hernández Flores
Facultad de Ciencias Políticas y Sociales, Universidad Nacional Autónoma de México (UNAM), Ciudad de México, México,
correo electrónico: armishi1122@gmail.com
Carlos Serrano Sánchez
Instituto de Investigaciones Antropológicas (IIA), Universidad Nacional Autónoma de México (UNAM), Ciudad de México, México,
correo electrónico: cserrano@unam.mx
Alejandro Terrazas Mata
Instituto de Investigaciones Antropológicas (IIA), Universidad Nacional
Autónoma de México (UNAM ),
correo electrónico: tema@unam.mx
Recibido el 17 de noviembre de 2021; aprobado el 16 de febrero de 2022
Resumen: El presente trabajo es un análisis de la variación morfológica de la mandíbula en restos prehistóricos de México, cuya cronología corresponde a la transición Pleistoceno tardío-Holoceno temprano y medio. Los restos proceden de distintos puntos del territorio mexicano: la cuenca de México, valle de Tehuacán, valle Puebla-Tlaxcala y de la península de Yucatán. La morfología de estos especímenes fue comparada con otras muestras mandibulares de periodos cronológicos más recientes, de los horizontes Preclásico y Clásico de Mesoamérica.
A partir del estudio de 13 medidas, se aplicaron un análisis univariado y dos multivariados. Los resultados sugieren que existe una alta variabilidad en los restos de las primeras poblaciones que ocuparon México.
Palabras clave: características mandibulares, poblaciones precerámicas, poblamiento antiguo, Altiplano Central, costa Caribe.
Abstract: The present work is an analysis of the morphological variation of the mandible in prehistoric remains from Mexico, whose chronology corresponds to the late Pleistocene-early and middle Holocene transition. The remains come from different regions of the Mexican territory: the basin of Mexico, the Tehuacan valley, the Puebla-Tlaxcala valley and the Yucatan Peninsula. The morphology of these specimens was compared with other mandibular samples from more recent chronological periods, the Preclassic and Classic horizons of Mesoamerica.
From the study of 13 measures, a univariate and two multivariate analyzes were performed. The results suggest a high variability in the remains of the first populations that occupied Mexico.
Key words: Mandibular osteometric variability, pre-ceramic population osteology, ancient settlement, Mexican central highlands, Yucatan Caribbean coast.
Introducción
El territorio que ocupa México en el continente americano, lo convierte en un sitio necesariamente de paso en el antiguo poblamiento humano de esta parte del mundo. América fue el último de los continentes en ser habitado por el Homo sapiens moderno y sea cual sea la ruta que siguió en su dispersión hacia su interior, es un hecho que cruzó por nuestro país. Prueba de ello son las evidencias de diversas expresiones culturales que han aparecido en distintos puntos del territorio, como son: artefactos líticos, huesos modificados, pinturas rupestres y algunos otros vestigios de ocupación humana en cuevas y abrigos rocosos. Así como la presencia misma, aunque escasa, de los restos esqueléticos pertenecientes a los primeros pobladores de México.
Los hallazgos de restos óseos humanos prehistóricos en nuestro país son pocos y algunos de ellos no han sido el resultado de proyectos destinados a su localización, sino más bien, producto de encuentros accidentales o fortuitos. Este hecho ha contribuido a la pérdida de algunos datos, como la ubicación exacta, la posición del esqueleto, la capa estratigráfica en que se encontró, la aparición o no de elementos asociados y quizá la pérdida misma de algún segmento del esqueleto o bien, de todo el esqueleto poscraneal, ya que, en muchos casos se ha recuperado solamente el cráneo junto con la mandíbula, en otros, sin ella y en algunos más, únicamente se cuenta con este último segmento óseo.
La mandíbula es una estructura ósea de gran importancia en los estudios de restos fósiles, ya que se trata de uno de los huesos más resistentes o que mejor perdura al paso del tiempo y en muchos casos, es lo único que se conserva (Loth y Henneberg, 1996; Kaifu y Baba, 2005; White et al., 2000; Rosas, 2001; Rosas y Aguirre, 1999; Walker et al., 2010). Por otro lado, se trata de un segmento móvil que se encuentra unido al cráneo a través de una articulación y al igual que él, permite plantear estudios de manera aislada, sobre variabilidad o afinidad poblacional a partir de diversos caracteres descriptivos y métricos (Morant, 1936; Albuquerque, 1952; Lagunas, 1967; Hernández y Martínez, 2008).
Estudios realizados en restos prehistóricos han registrado que durante el proceso evolutivo del hombre, la morfología del esqueleto poscraneal se ha mantenido relativamente constante, en cambio, es notoria una mayor diferenciación en las estructuras craneodentales, entre las que se encuentra estrechamente involucrada la mandíbula. Aquí radica su particular importancia desde el punto de vista de la prehistoria, así como de la antropología en general.
Diversos análisis llevados a cabo en el craneofacial de los restos óseos correspondientes a los primeros pobladores de América, han mostrado que existen al menos dos perfiles morfológicos distintos; uno de ellos cronológicamente más antiguo que data de más de 8000 años AP, presenta rasgos fenotípicos bien definidos, como es la forma alargada del cráneo y un prognatismo pronunciado. En cambio, se ha observado otro perfil con una cronología más tardía, el cual se caracteriza por una morfología más especializada (cráneos redondeados con caras planas, entre otros rasgos). Esta constatación dio lugar a un modelo de poblamiento que se conoce como “modelo de dos componentes” y ha sido ampliamente estudiado (Neves y Pucciarelli, 1989, 1991; Powell y Neves, 1999; Neves et al., 1999; Powell et al., 1999; González-José et al., 2001; Neves y Martínez, 2005; Neves et al., 2004; González-José et al., 2005; Jantz y Douglas, 2001; Pucciarelli, 2004, 2009; Pucciarelli et al., 2003; Hubbe et al., 2011).
En México, los estudios realizados en los restos esqueléticos de los primeros pobladores, han revelado una situación similar a lo antes expuesto; cráneos que superan los 9000 años AP se caracterizan por presentar formas elongadas, aquellos que se encuentran alrededor de los 7000 años AP tienden hacia la mesocránea y los más recientes, hacia el acortamiento del cráneo (Romano, 1955, 1963, 1970, 1974; Salas et al., 1988; Pompa y Serrano, 2001; Bautista y Pijoán, 2002; Jiménez et al., 2003, 2006, 2009, 2014; Jiménez y Hernández, 2011). En esta perspectiva surgió el presente trabajo, además de que los estudios que se han llevado a cabo en restos prehistóricos americanos se han abordado desde el análisis del craneofacial, excluyendo la mandíbula; ya que ésta tiene una estrecha relación con el cráneo, se consideró la posibilidad de encontrar ciertas características indicadoras de las afinidades biológicas entre los grupos involucrados en el primer poblamiento de América y su relación con poblaciones posteriores.
Para llevar a cabo esta investigación, se tuvo en cuenta la variación geográfica y cronológica de una muestra representativa del periodo Precerámico (cuenca de México, valle de Tehuacán, valle de Tulancingo y península de Yucatán). Estos datos fueron confrontados con los de otras muestras del periodo prehispánico (Tlatilco, de la cuenca de México; valle de Tehuacán; Chichen Itzá, del área maya). Se llevó a cabo después la evaluación de una serie de variables cuantitativas, que fueron sometidas a estadísticos univariados y multivariados para estimar las afinidades y diferencias entre los individuos que representan a cada una de las muestras empleadas en este estudio.
Este trabajo pretende contribuir al conocimiento de la población prehistórica de México y del continente americano, pues no sólo se trata de los primeros pobladores de nuestro país, sino que también forman parte de los primeros habitantes de América.
Material y método
La muestra está conformada por especímenes que forman parte de la colección de restos óseos humanos precerámicos de México, resguardados en la Dirección de Antropología Física (DAF) del Instituto Nacional de Antropología e Historia (INAH), y un ejemplar preservado en el Laboratorio de Prehistoria y Evolución Humana (LPEH) del Instituto de Investigaciones Antropológicas de la Universidad Nacional Autónoma de México (UNAM). Estos materiales están considerados cronológicamente como los más antiguos de México y de acuerdo con los fechamientos practicados a algunos de ellos,1 como se puede ver en la Tabla 1, el margen de antigüedad de la muestra oscila entre los 11000 y los 4000 años AP.
Como se ha mencionado, la mayoría de ellos fueron localizados en la Ciudad de México y el Estado de México, otros más en Puebla, Hidalgo y Quintana Roo, como se muestra en la Figura 1.
Tabla 1. Esqueletos precerámicos que conforman la muestra (fechamientos sin calibrar)
|
Espécimen |
Código |
Sexo |
Fechamiento |
Referencia |
|
Balderas |
BALD |
Masculino |
10 500 AP |
González et al., 2003 |
|
Santa María Astahuacán II |
ASTAH2 |
Masculino |
10 300 + 600 AP |
Berger y Protsch, 1989 |
|
Santa María Astahuacán III |
ASTAH3 |
Femenino |
10 300 + 600 AP |
Berger y Protsch, 1989 |
|
Peñón III |
PEÑIII |
Femenino |
10 755 + 75 AP |
González et al., 2003 |
|
Chimalhuacán |
CHIMAL |
Masculino |
10 500 AP |
González et al., 2003 |
|
Texcoco* |
TEXC |
Masculino |
10 000-6 000 AP |
Morett, 2004 |
|
Texcal (Ent. 3 múltiple, suj. 1) |
TEXCA |
Masculino |
7 480 + 55 AP |
González et al., 2003 |
|
Texcal (Ent. 3 múltiple, suj. 3)* |
TEXCA3 |
Femenino |
7000 - 4 500 AP |
García Moll, 1977 |
|
Texcal I |
TEXCA1 |
Femenino |
7 233 + 36 AP |
Jiménez y Hernández, 2011 |
|
Tehuacan, ent. 2 TC 272 |
TEH2 |
Masculino |
4 121 + 96 AP |
Johnson y MacNeish, 1972 |
|
Tehuacan, ent. 3 TC 272 |
TEH3 |
Femenino |
4 121 + 96 AP |
Johnson y MacNeish, 1972 |
|
Tehuacan, ent. 4 TC 50 |
TEH4 |
Masculino |
6 513 + 186 AP |
Johnson y MacNeish, 1972 |
|
Cuenca de México |
CUMEX2 |
Masculino |
3 024 + 48 AP |
Jiménez et al., 2016 |
|
Peñón del Marqués I |
PMARQ2 |
Masculino |
4 247 + 29 AP |
Jiménez et al., 2016 |
|
Tecolote B* |
TECOLB |
Masculino |
9000-7 000 AP |
Lorenzo, 1967 |
|
Las Palmas |
PALM |
Femenino |
8 050 + 130 AP |
González et al., 2008 |
Nota: * fechamientos estimados.
Figura 1. En color gris se indican las regiones en donde se han localizado restos humanos prehistóricos
Las mandíbulas que se utilizaron en esta investigación pertenecen a individuos adultos, masculinos y femeninos. La estimación de la edad y el sexo se realizó siguiendo diferentes parámetros (Ubelaker, 1978; Meindl y Lovejoy, 1985; Lovejoy et al., 1985; White y Folkens, 2005). En el caso específico de la mandíbula, estructura que también presenta características sexuales importantes, además de considerar el tamaño y la robustez, se evaluaron otras características dimórficas que se concentran principalmente en la rama ascendente y en la región del mentón (Loth y Hennenberg, 1996; Coquerelle
et al., 2011).
Análisis estadístico
Se aplicaron tres tipos de pruebas: un análisis univariado y dos multivariados. Estos últimos permiten analizar de forma simultánea diversas variables, para obtener un mejor entendimiento del fenómeno que se está estudiando (Härdle y Simar, 2007).
El análisis univariado consideró cada una de las medidas por separado. Para ello, se utilizó la distribución de frecuencias y las medidas de tendencia central de cada variable. En cambio, para los análisis multivariados se usaron dos tipos de pruebas; la primera de ellas fue el análisis de componentes principales (PCA, por sus siglas en inglés), la cual reduce la dimensionalidad de un conjunto de datos, busca las causas de la variabilidad de dichos datos y las ordena por importancia, donde la varianza de mayor tamaño es capturada en el primer eje o componente principal y así sucesivamente, según se vaya reduciendo la varianza. Es importante mencionar que este procedimiento de análisis se considera libre de hipótesis, por lo cual, los datos no requieren supuestos de distribución previos y son resistentes a los tamaños muestrales.
El segundo análisis multivariado consistió en un análisis discriminante, el cual permite conocer o buscar, si es que existen, las diferencias significativas entre grupos, a partir de una serie de variables, y determina cuál de ellas explica la mayor parte de las diferencias observadas. Finalmente, anotamos que para la aplicación de todas las pruebas estadísticas, se utilizó el programa estadístico Past, el cual fue destinado inicialmente para trabajos en paleontología, pero es también de gran utilidad en este tipo de investigaciones.
Resultados
Para el análisis se utilizaron 13 medidas (véase Tabla 2), todas ellas tomadas en el lado izquierdo; no obstante, debido a que los materiales, en su mayoría, no se encuentran en buen estado de conservación y hay zonas faltantes, se realizó un procedimiento de imputación o reemplazo de valores perdidos; lo cual se estableció de acuerdo a dos criterios: el primero fue asumir simetría, puesto que las variables que fueron analizadas son bilaterales, el lado considerado fue el izquierdo y cuando éste no se presentó fue sustituido por el valor del lado derecho. El segundo criterio consistió en actualizar aquellos datos, cuyo sujeto tuviera menos del 5% de datos perdidos, por el valor equivalente de su vecino más cercano (neighbour joining), para lo cual se utilizó la matriz de distancias en los casos en que no se cumplía con dichos requisitos.
Tabla 2. Variables utilizadas
|
Código |
Medida |
|
ANMR |
Anchura máxima de la rama |
|
ANMIR |
Anchura mínima de la rama |
|
ALPR |
Altura posterior de la rama |
|
ALMIR |
Altura mínima de la rama |
|
ALAR |
Altura anterior de la rama |
|
ALSIG |
Altura o profundidad de la escotadura sigmoidea |
|
ANSIG |
Anchura de la escotadura sigmoidea |
|
APCOD |
Diámetro antero-post del cóndilo |
|
TCOD |
Diámetro transverso del cóndilo |
|
GCM |
Grosor del cuerpo entre M1 y M2 |
|
ALCM |
Altura del cuerpo entre M1 y M2 |
|
LDCM |
Longitud directa del cuerpo |
|
ANGM |
Ángulo de la mandíbula |
Análisis univariado
En este análisis se estimó la media y la mediana de cada una de las variables, para posteriormente ser comparadas por población, entre masculinos y femeninos de los Precerámico y del Preclásico (Tabla 3 y Tabla 4). En la primera de ellas (la media), observamos que para la anchura máxima de la rama (ANMR), el valor más alto lo ocupan los individuos precerámicos, en cambio en los femeninos del Clásico se presenta el valor más bajo, lo cual indica que se trata de ramas más estrechas en la región superior.
En el caso de la anchura mínima de la rama (ANMIR) se observa que los valores de los grupos precerámicos, tanto femeninos como masculinos, son los que mantienen los valores más altos, sin embargo, es muy similar a los grupos del Preclásico, en tanto que las mujeres del Clásico son las que presentan una mayor estrechez en esta zona de la rama. En la altura posterior de la rama (ALPR), el grupo precerámico es el que posee los valores más altos y en el caso de los femeninos del Clásico, son los que tienen el valor notoriamente más bajo.
En la altura mínima de la rama (ALMIR) nuevamente el valor más alto se encuentra en sujetos masculinos del grupo precerámico, en cambio, en los femeninos sucede lo contrario, al poseer las mujeres del Preclásico el valor más alto. De forma similar se comporta la media en la ALAR que también valora la altura de la rama, sólo que en este caso se refiere a la región anterior.
Para la ALSIG que describe la profundidad de la escotadura sigmoidea, nuevamente el valor más alto de la media se encuentra en los masculinos del periodo Precerámico, por el contrario, en los grupos del Preclásico su valor es muy similar para ambos sexos, siendo incluso ligeramente más alto en los femeninos. En la ANSIG (anchura de la escotadura sigmoidea) los valores más altos los poseen en general, los individuos de sexo masculino.
El diámetro transverso del cóndilo (TCOD) muestra valores homogéneos en todos los grupos, ligeramente más alto en los masculinos del Precerámico y más bajo en los femeninos del Clásico. Una distribución similar se observa en el diámetro anteroposterior del cóndilo (APCOD). En cuanto al grosor del cuerpo de la mandíbula entre el primer y segundo molar (GCM), fue el grupo del Preclásico (tanto masculinos como femeninos) el que mantiene el valor más alto de la media, esto es, poseen un mayor grosor del cuerpo, mientras que el más bajo se encuentra en los femeninos del periodo Precerámico, al ser más estrechos. En la altura del cuerpo de la mandíbula entre M1 y M2 (ALCM) son los masculinos del Preclásico quienes poseen el valor más alto, es decir, presentan cuerpos con mayor altura en esta zona.
En lo que respecta a la longitud directa del cuerpo de la mandíbula (LDCM) los masculinos del periodo Precerámico cuentan con el valor más alto, esto es, son de mandíbulas más largas, a diferencia de los femeninos de este mismo periodo, que poseen el valor más bajo al ser más cortas, mientras que las del Preclásico y Clásico presentan medias muy similares. Finalmente, en el ángulo de la mandíbula (ANGM), son las mujeres del Clásico las que poseen la media más alta, lo que nos refiere ángulos más abiertos y por lo contrario, los masculinos de la etapa precerámica, la más baja, al tener ángulos más cerrados.
En términos generales, los valores más altos de la media de casi todas las variables, se encuentran en los individuos masculinos y particularmente, en los del periodo Precerámico. En cambio, los más bajos se ubican principalmente en los femeninos del Clásico, a excepción de ANGM, donde sucedió lo contrario. Mientras que el grupo del Preclásico mostró un mayor número de variables con valores intermedios, lo cual se observa más claramente en los grupos femeninos.
Tabla 3. Estadística descriptiva que compara la distribución de los valores de la media de todas las variables, entre todos los grupos
|
|
Comparación de medias |
|||||||||||||
|
|
|
ANMR |
ANMIR |
ALPR |
ALMIR |
ALAR |
ALSIG |
ANSIG |
TCOD |
APCOD |
GCM |
ALCM |
LDCM |
ANGM |
|
Masc. |
Precerámico |
48.50 |
36.36 |
64.45 |
52.18 |
67.82 |
15.45 |
38.55 |
22.91 |
11.41 |
16.05 |
31.41 |
93.43 |
122.86 |
|
Preclásico |
46.74 |
35.82 |
58.74 |
51.89 |
65.84 |
13.24 |
37.08 |
21.42 |
11.18 |
17.11 |
32.18 |
90.61 |
126.55 |
|
|
Fem. |
Precerámico |
47.00 |
35.42 |
57.83 |
45.92 |
57.75 |
12.58 |
38.00 |
21.25 |
10.92 |
14.50 |
28.00 |
87.67 |
124.58 |
|
Preclásico |
44.93 |
34.93 |
52.00 |
47.57 |
62.14 |
13.71 |
36.14 |
19.86 |
11.07 |
15.86 |
29.36 |
88.93 |
128.00 |
|
|
Clásico |
43.1 |
32.5 |
46.5 |
41.8 |
54.2 |
11.8 |
35.2 |
19.4 |
9.7 |
16.1 |
24.9 |
88.2 |
129.3 |
|
Tabla 4. Estadística descriptiva que compara la distribución de los valores de la mediana de todas las variables, entre todos los grupos
|
|
Comparación de la mediana |
|||||||||||||
|
|
|
ANMR |
ANMIR |
ALPR |
ALMIR |
ALAR |
ALSIG |
ANSIG |
TCOD |
APCOD |
GCM |
ALCM |
LDCM |
ANGM |
|
Masc. |
Precerámico |
49 |
36 |
62 |
52 |
67 |
15 |
38 |
22.5 |
11.5 |
16 |
31 |
93 |
124 |
|
Preclásico |
47 |
36 |
58 |
53 |
66 |
13 |
37 |
21 |
11 |
17 |
33 |
90.5 |
126 |
|
|
Fem. |
Precerámico |
46.5 |
35.5 |
59 |
44.75 |
58 |
12.25 |
37.75 |
21.5 |
10.5 |
14 |
30 |
87 |
125.25 |
|
Preclásico |
44.5 |
35.5 |
51 |
46 |
60 |
13.5 |
37 |
19 |
12 |
16 |
30 |
89 |
130 |
|
|
Clásico |
43.5 |
34 |
47 |
41 |
56 |
12 |
35 |
20.5 |
9 |
16 |
24 |
87 |
128 |
|
Análisis de componentes principales
El análisis de componentes principales (PCA) reunió el 83.80% de la varianza de las variables originales con las tres primeras componentes, lo cual se considera suficiente para el estudio. En la PC1, el 59.12% explicó las variables correspondientes a las alturas posterior (ALPR), mínima (ALMIR) y anterior (ALAR) de la rama ascendente en sus valores positivos y la variable del ángulo de la mandíbula (ANGM) hacia sus valores negativos. En la PC2, el 14.57% explicó las variables que refieren a la anchura máxima de la rama ascendente (ANMR), anchura de la escotadura sigmoidea (ANSIG), longitud directa del cuerpo de la mandíbula (LDCM) y ángulo de la mandíbula (ANGM), todas ellas hacia sus valores positivos. Por último, la PC3 explicó el 10.11% de las variables altura mínima (ALMIR) y anterior (ALAR) de la rama ascendente, altura del cuerpo (ANSIG) y el ángulo de la mandíbula (ANGM) hacia sus valores positivos, y la anchura máxima de la rama ascendente (ANMR) junto con la anchura de la escotadura sigmoidea (ANSIG) en sus valores negativos.
En términos generales, la primera componente principal (PC1) describió la variación a partir del sexo, es decir, los individuos femeninos se ubican principamente hacia los valores negativos, mientras que los masculinos se ubican en los positivos. No obstante, se observó amplio traslape hacia el centro. Mientras que las PC2 y PC3, pueden ser entendidas como componentes con una variabilidad principamente aleatoria.
Como es posible apreciar en la Figura 2, los individuos masculinos de la época precerámica se ubican mayoritariamente hacia los valores positivos de la PC1 y en el rango completo de la PC2. Lo anterior, en términos morfológicos, implica que estos sujetos poseen cualidades morfológicas que reflejan robustez de la rama mandibular. Por su parte, es posible observar que los individuos masculinos del Preclásico, comparten mayoritariamente dichas cualidades, no obstante, se aprecia un conjunto de éstos que se ubican hacia los valores negativos de la PC1, lo cual tendría que ser interpretado como una mayor variabilidad al interior de este grupo, para estas características de la robustez de la rama mandibular.
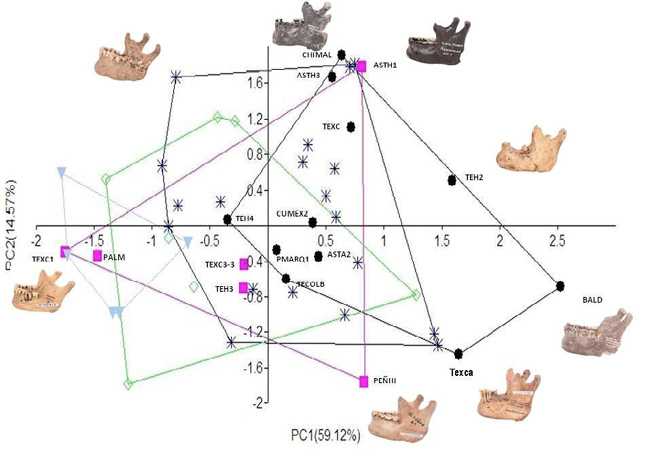
Figura 2. Gráfica de dispersión de las dos primeras componentes principales. (●) círculo negro= masculinos, Precerámico; (■) cuadro rosa= femeninos, Precerámico; (*) azul intenso= masculinos, Preclásico; (◊) rombo verde= femeninos, Preclásico, y (▼) triángulo azul claro= femeninos, Clásico
En cuanto al grupo femenino del periodo Precerámico, es posible ver que la mayoría se ubica hacia los valores negativos de la PC1 y en cambio, dos de ellas se encuentran en los valores positivos, se trata de la Mujer del Penón III y la Mujer de Astahuacan I. Ambos casos coiciden en cronología, pues poseen los fechamientos más antiguos de toda la muestra y por otro lado, mostrarían lo que podemos considerar rasgos de perfil más masculino, a diferencia del resto de los sujetos femeninos. Este rasgo concuerda con lo que Enlow (1990) dice acerca de las características dimórficas relacionadas con el sexo de los sujetos femeninos con cráneos cortos y alargados. Dicha distinción radica en que la braquicefalia femenina aumenta o resalta las características propias de su sexo y en cambio, la mujer dolicocéfala, al presentar rasgos faciales más estrechos y protrusivos, tiende a presentar un caracter más masculino en la cara. En este caso, ambos sujetos se encuentran dentro de la clasificación de dolicocráneos.
En este análisis, se puede ver que estos dos ejemplares poseen cierta robustez de la rama mandibular, semejante a lo que ocurre con los individuos del sexo masculino, mientras que el resto de las mujeres de la época precerámica se caracterizan por ser de baja altura y poca anchura de la rama mandibular.
En el caso de las poblaciones femeninas del Preclásico y Clásico, también se ubican dentro de los valores negativos, que como se ha mencionado, están relacionados con la variabilidad de la rama mandibular. Particularmente, los ejemplos de Chichen-Itzá, que corresponden al Clásico, puede notarse que se ubican en los valores más extremos negativos de la PC1, adicionalmente, se observa que son una agrupación sólida y con cierta homogeneidad.
Como se puede ver, la PC1 separó a los individuos mayoritariamente debido al sexo, es decir, masculinos hacia los valores positivos y femeninos en los negativos. A su vez, se ha mencionado que a lo largo de esta componente se resumen las variables de la rama mandibular (alturas y anchuras de diferentes regiones de la rama). Esto puede ser entendido como indicador del dimorfismo sexual en la estructura de la mandíbula, lo cual coincide con estudios previos que han sido realizados en este segmento óseo (Lagunas, 1967; Hernández y Martínez, 2008).
Como se observa en la Figura 3, la PC2 presenta una distribución similar a la PC1 para los tres grupos femeninos, esto es, se distribuyen mayormente en los valores negativos, lo cual es aún más acentuado en el grupo del Clásico. Cabe mencionar que en este componente, al igual que en la PC1, los casos especificos de la Mujer del Peñón III y Astahuacán I nuevamente se diferencían del resto del grupo precerámico, manteniendo una mayor cercanía entre ellas en valores positivos de la PC2. Lo que en términos morfológicos está indicando que ambos individuos presentan una mayor robustez en la rama ascendente de la mandíbula. En cambio, el resto de los individuos del periodo Precerámico, Preclásico y Clásico, presentan una mayor semejanza entre ellos, siendo aún más graciles las mujeres del Clásico.

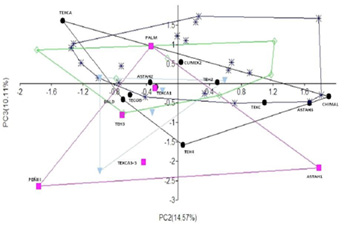
Figura 3. Gráfico de dispersión de las tres primeras componentes principales. Se contrastan la PC1 vs PC3 (izquierda) y PC2 vs PC3 (derecha). (●) círculo negro= masculinos, Precerámico; (■) cuadro rosa= femeninos, Precerámico; (*) azul fuerte= masculinos, Preclásico; (◊) rombo verde= femeninos, Preclásico, y (▼) triángulo azul claro= femeninos, Clásico
En el caso de la PC3, se observa un traslape importante entre los grupos, aunque los ejemplos femeninos de la época precerámica muestra cierta tendencia a segregarse del resto de las poblaciones.
Análisis discriminante
Con el propósito de describir sí existen diferencias entre grupos cuando las variables de estudio son analizadas de manera conjunta se utilizó el análisis discriminante. Este análisis basado en la matriz de distancias de Mahalanobis, nos permite verificar la consistencia de los K-grupos y las reclasificaciones de los casos. De esta manera, se entiende que las nuevas variables canónicas (CV) son una función lineal con que se logra maximizar las diferencias entre grupos. Este abordaje permite inferir semejanzas o diferencias entre grupos.
En este análisis el valor del lambda de Wilks mide las desviaciones de las puntuaciones discriminantes dentro de los grupos respecto a las desviaciones totales, sin distinguir grupos. Así, si su valor tiende a uno, la dispersión es debida a que no existen diferencias entre grupos, dicho de otra manera, es la proporción de la varianza total en las puntuaciones discriminantes que no es explicada por la diferencia entre grupos. Valores cercanos a cero indican que los grupos son diferentes.
En el presente análisis, siguiendo la metodología de Ackerman (2006, p. 636), se ha removido el efecto de la variación debida al sexo. De esta manera, el estudio comparativo multivariado, para evaluar diferencias entre grupos, puede ser realizado sobre la base de las diferentes cronologías. Se considera que esta aproximación es adecuada debido a que, como se vio en el análisis exploratorio, las diferencias sexuales de la mandíbula constituyen la principal fuente de variación y esto podría conducir a falsas interpretaciones respecto a la diversidad existente entre grupos. No obstante, decidimos conservar el efecto del tamaño debido a que ciertas características responden a efectos estocásticos y adaptativos (Rodríguez, 2001; Pucciarelli, 2004; Mirazon-Lahr, 1995).
Con lo anterior, ha sido posible evaluar que las diferencias entre grupos, cuando se analizan de manera multivariada, no son significativas (sig.= 0.106). No obstante, el estadístico lambda de Wilks mostró un valor de LW=0.405, lo cual indica que existe una alta variabilidad al interior de los grupos en relación a la que existe entre grupos. Esto puede ser verificado en la Tabla 5, que se refiere a la validación de las clasificaciones, donde se muestra que un 70.8% de los casos se clasificaron correctamente, dicho de otra manera, tres de cada diez ejemplos se reclasificaron en un grupo diferente del original. No obstante, en la validación cruzada por la permutación Jacknifed sólo el 43.8% de los casos se clasificaron de acuerdo a su grupo de origen.
Tabla 5. Matriz de clasificación en el análisis discriminante
|
|
|
Precerámico |
Preclásico |
Clásico |
Total |
|
Reclasificación |
|||||
|
n |
Precerámico |
11 |
4 |
2 |
17 |
|
|
Preclásico |
4 |
19 |
3 |
26 |
|
|
Clásico |
0 |
1 |
4 |
5 |
|
% |
Precerámico |
64.71 |
23.53 |
11.76 |
100 |
|
|
Preclásico |
15.38 |
73.08 |
11.54 |
100 |
|
|
Clásico |
0 |
20 |
80 |
100 |
|
Validación cruzada |
|||||
|
n |
Precerámico |
9 |
6 |
2 |
17 |
|
|
Preclásico |
9 |
11 |
6 |
26 |
|
|
Clásico |
2 |
2 |
1 |
5 |
|
% |
Precerámico |
52.94 |
35.29 |
11.76 |
100 |
|
|
Preclásico |
34.62 |
42.31 |
23.08 |
100 |
|
|
Clásico |
40 |
40 |
20 |
100 |
Clasificados correctamente el 70.8% de los casos agrupados originales.
Clasificados correctamente el 43.8% de los casos agrupados mediante validación cruzada.
Como se observa en la Figura 4, entre las poblaciones del Precerámico y el Preclásico existe un amplio traslape debido a la diversidad dentro de grupos; a su vez, son los ejemplos del Clásico los que se encuentran mayormente separados. Esto, conjuntamente con los resultados del lambda de Wilks, nos permite observar que dentro de cada agrupación hay mucha variabilidad de la forma de la mandíbula; lo cual es sencillo de entender, considerando que para el caso de los precerámicos se abarca un periodo cronológico de aproximadamente ocho mil años y de igual manera, en el caso del Preclásico existe cierta heterogeneidad debida a la cronología. Adicionalmente a ello, hay que considerar que la metodología de Ackerman (2006) remueve los efectos de la tendencia central, no obstante, se conservan las varianzas de grupos, es decir, se adhieren las varianzas de cada uno de los sexos en las diferentes agrupaciones. Si bien esto puede ser entendido como un factor que suaviza las diferencias entre grupos, considerando que el análisis discriminante logra resolver el vector de mayor separación de grupos, es posible tomar en cuenta que este resultado guarda mayor congruencia que si se analizaran los sexos de manera separada. Esto se ve reforzado considerando que los sujetos de mayor antigüedad, se ubican hacia los valores negativos de la primera raíz canónica (CV1) y se observa cierta tendencia en dirección hacia los positivos de la misma, donde se ubican los ejemplares del Preclásico y Clásico.
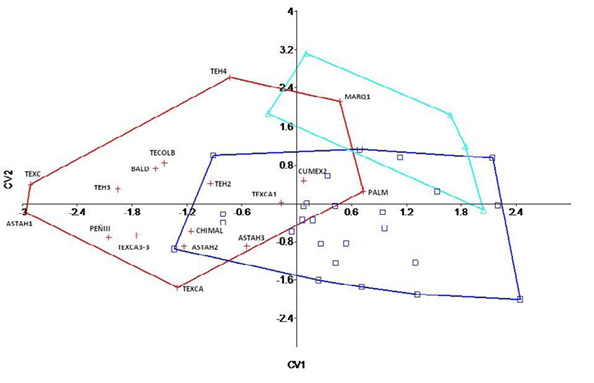
Figura 4. Gráfico de dispersión de la primera (CV1) y segunda (CV2) raíces canónicas discriminantes. (+) Cruz roja= Precerámico, (□) cuadro azul= Preclásico y (▲) triángulo turquesa= Clásico
Como se observa en la Figura 4, entre las poblaciones del Precerámico y el Preclásico existe un amplio traslape debido a la diversidad dentro de grupos; a su vez, son los ejemplos del Clásico los que se encuentran mayormente separados. Esto, conjuntamente con los resultados del lambda de Wilks, nos permite observar que dentro de cada agrupación hay mucha variabilidad de la forma de la mandíbula; lo cual es sencillo de entender, considerando que para el caso de los precerámicos se abarca un periodo cronológico de aproximadamente ocho mil años y de igual manera, en el caso del Preclásico existe cierta heterogeneidad debida a la cronología. Adicionalmente a ello, hay que considerar que la metodología de Ackerman (2006) remueve los efectos de la tendencia central, no obstante, se conservan las varianzas de grupos, es decir, se adhieren las varianzas de cada uno de los sexos en las diferentes agrupaciones. Si bien esto puede ser entendido como un factor que suaviza las diferencias entre grupos, considerando que el análisis discriminante logra resolver el vector de mayor separación de grupos, es posible tomar en cuenta que este resultado guarda mayor congruencia que si se analizaran los sexos de manera separada. Esto se ve reforzado considerando que los sujetos de mayor antigüedad, se ubican hacia los valores negativos de la primera raíz canónica (CV1) y se observa cierta tendencia en dirección hacia los positivos de la misma, donde se ubican los ejemplares del Preclásico y Clásico.
Mediante el análisis discriminante, no es posible realizar la inferencia de la antigüedad de un ejemplar. Sobre todo, debido a que se analiza la variabilidad dentro de una misma especie, donde las diferencias suelen no ser tan grandes. No obstante, las reclasificaciones de los sujetos no representan una negación de su correspondencia a cierta cronología, como por ejemplo el sujeto de Las Palmas, datado 8050 + 130 AP por la técnica de C14, sino que permiten ubicar sujetos informativos sobre la amplitud de la variabilidad en una cronología específica. Esto es una ventaja, considerando que aunque no se corresponden en cuanto a fenotipo y su datación absoluta es conocida, el análisis discriminante nos indica que dentro de cada una de las poblaciones la variabilidad morfológica de la mandíbula es muy amplia, traslapando incluso las barreras de la temporalidad.
Discusión
Hasta ahora, estudios realizados en la morfología del cráneo en la población prehistórica de México, han indicado que la variación craneofacial va más allá de los conceptos paleoamericano y amerindio, por el contrario, se ha visto que existe una amplia variabilidad en esta población (Terrazas y Benavente, 2006; Chatters et al., 2014; Serrano et al., 2016; Hernández, 2018). Los resultados obtenidos en este trabajo así lo reafirman; la muestra de mandíbulas del periodo Precerámico, presentó una alta variabilidad a su interior al igual de lo que se aprecia en el craneofacial.
Al encontrar estas similitudes entre el cráneo y su respectiva mandíbula (en estudios por separado), se optó por calcular el índice mandibular esperando encontrar semejanzas en la forma de ambas estructuras, puesto que las variaciones en la forma de la cabeza determinan los cambios correspondientes en el tipo y patrón faciales (Enlow, 1990). No obstante, fueron pocos los casos que presentan cierta armonía entre la forma del cráneo y la forma de la mandíbula.2 Al calcular los índices craneal horizontal máximo y mandíbula3 varios de ellos no muestran esta concordancia, puesto que se esperaría que un cráneo largo tuviera una mandíbula larga y como se puede observar en la Tabla 6, esto no sucedió así.
Al respecto, Albuquerque (1952) realizó un trabajo similar en una población portuguesa, ella menciona que el índice mandibular concordó con el índice craneal, que en este caso se trató de un grupo predominantemente dolicocéfalo. Por su parte Lagunas (1967), llevó a cabo un estudio en mandíbulas prehispánicas de la población tlatelolca y encontró que en ellas predomina la forma braquignata y en menor porcentaje las mesognatas, sin embargo, a pesar de que él no midió los respectivos cráneos de las mandíbulas, retoma los datos aportados por Dávalos (1951) quién estudio la misma población y reportó que un mayor porcentaje fue de cráneos cortos, seguido por los de forma media. Por lo que Lagunas consideró que, en cierta medida, el índice mandibular corresponde con el craneal.
Tabla 6. Contrastación de los índices craneal horizontal máximo y mandibular
|
Esqueleto |
Mandibular |
Craneal |
Cronología |
|
Peñón III |
Braquignata |
Dolicocráneo |
10 755 + AP |
|
Balderas |
Dolicognata |
Dolicocráneo |
10 500 AP |
|
Chimalhuacán |
- |
Dolicocráneo |
10 500 AP |
|
Astahuacán 1 |
Braquignata |
Dolicocráneo |
10 300 + 600 AP |
|
Astahuacan 2 |
- |
Mesocráneo |
10 300 + 600 AP |
|
Astahuacan 3 |
Mesognata |
- |
10 300 + 600 AP |
|
Texcoco |
- |
Dolicocráneo |
10 000–6000 AP |
|
Palm |
- |
Mesocráneo |
8 050 + 130 AP |
|
Tecolote B |
Braquignata |
Dolicocráneo |
9000 - 7000 AP |
|
Texcal 3-3 mult. |
Mesognata |
Dolicocráneo |
7 000 - 5000 AP |
|
Texcal 1 |
Mesognata |
Mesocráneo |
7 233 + 36 AP |
|
Texcal 3 mult. |
Braquignata |
Braquicráneo |
7 480 + 55 AP |
|
Tepexpan |
- |
Mesocráneo |
7 400 - 4 700 AP |
|
Coxcatlan 4 |
Mesognata |
Mesocráneo |
6513 + 186 AP |
|
Purrón 2 |
- |
- |
4121 + 96 AP |
|
Purrón 3 |
- |
- |
4121 + 96 AP |
|
Cuenca Mex. |
Dolicognata |
- |
3024 + 48 AP |
|
Peñón Marqués 1 |
Braquignata |
- |
4 247 + 29 AP |
Sin embargo, en este trabajo se registra que es clara la falta de relación entre la forma del neurocráneo y la mandíbula, sobre todo, en aquellos casos donde el cráneo es dolicoide y su respectiva mandíbula es braquignata (Figura 5 y Figura 6), como se presenta en la Mujer del Peñón III, Astahuacán I y el entierro B de la Cueva del Tecolote. Una forma de explicar este escenario podría ser a través de los conceptos de modularidad e integración morfológica, los cuales hacen referencia al grado de covariación entre los componentes de una estructura. Los módulos son estructuras que covarían fuertemente, pero a su vez son relativamente independientes unos de otros, mientras que la integración morfológica se refiere a la variación morfológica coordinada de los componentes de un todo funcional (Püschel, 2014). A partir de estos conceptos se plantea que la región facial, el neurocráneo y la base craneal son módulos que varían de manera semi-independiente, ya que estas estructuras interactúan de manera integral, donde cada unidad puede tener influencias desiguales entre unos y otros, debido a que cada módulo tiene una trayectoria de crecimiento distinta (Lieberman, 2008, 2011; Lieberman et al., 2002; Martínez-Abadías et al., 2011).
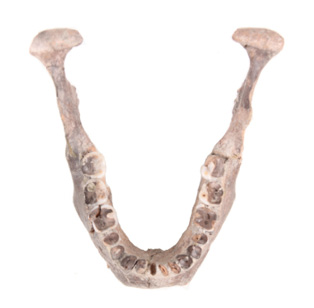
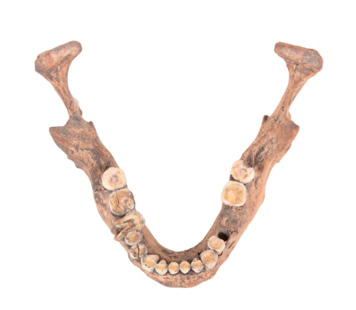
Figura 5. A la izquierda mandíbula dolicognata del Hombre de Balderas y a la derecha, mandíbula braquignata del entierro B de la Cueva del Tecolote. Ambos casos corresponden a individuos de sexo masculino con cráneo dolicoide. Fotografías: Leonardo Hernández Vidal
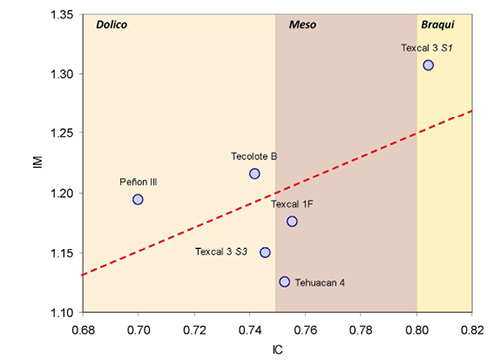
Figura 6. Relación entre el índice cefálico (IC) y el mandibular (IM) de individuos precerámicos de México. En términos generales se observa que existe cierta correspondencia entre la longitud y anchura craneal con respecto de la longitud y anchura de la mandíbula, no obstante, la correlación es baja y no significativa (r= 0.552; p= 0.256)
Lieberman (2011) menciona que el tamaño y forma del cerebro influye directamente en la forma del craneofacial sólo hasta que el crecimiento del cerebro se completa, lo cual ocurre o coincide con la aparición de los primeros molares permanentes. En cambio, la región facial tiene una trayectoria de crecimiento más lenta y prolongada, que puede ir más allá de la erupción de los terceros molares; y debido a que nervios y vasos sanguíneos que conectan el cerebro con el resto del cuerpo cruzan la base del cráneo, éste tiene una trayectoria de crecimiento más compleja.
La base del cráneo actúa como un integrador de la cabeza entre el cerebro y la cara, los cuales a su vez interactúan entre sí a través de la base craneal; sin embargo, estas interacciones no son iguales en términos de fuerza y sincronización. Los huesos que conforman la base craneal4 contribuyen a la formación de tres concavidades en esta zona: la fosa craneal posterior (PCF), media (MCF) y anterior (ACF). La primera de ellas alberga la parte posterior del cerebro (cerebelo y tallo cerebral), en la segunda subyacen los lóbulos temporales, la fosa hipofisaria y en la superficie inferior se establecen las posiciones de las cavidades glenoideas para la mandíbula y suturas craneofaciales para el complejo nasomaxilar; por tanto, en consecuencia, es una guía que también regula la anchura de la cara en el crecimiento (Enlow, 1990 y Lieberman, 2011).
En términos generales, Lieberman (2011) menciona que la base del cráneo crece y cambia de varias formas importantes, en donde las tres fosas se alargan en sentido anteroposterior y se amplían mediolateralmente. Su anchura, tiene una influencia importante en la forma craneofacial y viceversa. Sin embargo, debido a que la región facial crece a un ritmo diferente de las demás regiones de la cabeza, se considera que la integración entre la cara y el resto del cráneo sea probablemente desigual (Figura 6).
Conclusión
A pesar de que existe una carencia de estudios en la mandíbula de los primeros pobladores de América, los resultados de este trabajo permitieron registrar altos niveles de variación en las primeras poblaciones que ocuparon México. La diversidad biológica observada en algunas mandíbulas de los cráneos de los restos precerámicos muestra cierta tendencia espacial y temporal que no puede ser explicada a partir del modelo de “dos componentes” (Neves y Pucciarelli, 1989; 1991), por el contrario, esto nos hace suponer que los grupos prehistóricos experimentaron la influencia in situ de diferentes fuerzas microevolutivas durante el poblamiento temprano.
La mandíbula de Las Palmas es muy diferente de sus contemporáneas (pequeña y grácil), posiblemente por una reacción (adaptación) a las condiciones tropicales donde vivió (poblaciones de una especie que viven en el trópico tienden a ser de menor talla).
Finalmente, la presencia de una determinada forma del cráneo (dolico, meso o braqui), no determina la forma del craneofacial, incluyendo la mandíbula. Como se pudo observar en este trabajo, existe una amplia diversidad de fenotipos, en la cual algunas de las formas del craneofacial responden a la forma del neurocráneo y en algunos casos, esta relación es más compleja.
Agradecimientos
Al antropólogo J. Concepción Jimenez de la Dirección de Antropología Física del INAH, por las facilidades otorgadas para la revisión de los materiales y al doctor Jorge A. Gómez-Valdés, del Posgrado en Antropología Física, de la Escuela Nacional de Antropología e Historia, por su ayuda en el análisis estadístico. Al Programa de Apoyos a Proyectos de Investigación e Innovación Tecnológica (PAPIIT), UNAM, proyecto IN302219, “Historia biológica y dinámica poblacional en el México prehispánico. Una contribución antropológica, Instituto de Investigaciones Antropológicas (UNAM).
Referencias
Albuquerque, Rolanda M.
(1952) Estudo antropológico da mandíbula nos Portugueses. Contribuiçoes para o Estudo da Antropología Portuguesa, 5, 65-196.
Ackermann, R. R., Jeffrey R. y Cheverud, J. M.
(2006) Identifying the morphological signatures of hibridation in primate and human evolution. Journal of Human Evolution, 51, 632-645.
Bautista, Josefina y Pijoan, C. M.
(2002) Estudio métrico comparativo entre cráneos mexicanos (precerámicos y principios del siglo XX). Paleopatología, 50-58.
Berger, Rainer y Protsch, Reiner
(1989) UNCLA radiocarbon dates XI. Radiocarbon, 31 (1), 55-67.
Chatters, J.C., Kennett, D. J., Asmerom, Y., Kemp, B. M., Polyak, V., Nava, A., Beddows, P. A., Reinhardt, E., Arroyo-Cabrales, J., Bolnick, D. A., Malhi, R. S., Culleton, B. J., Luna, P., Rossolo, D., Morell-Hart, S. y Stanfford Jr., T. W.
(2014) Late Pleistocene human skeleton and mtDNA link paleoamericans and modern native Americans. Science, 344, 750-754.
Coquerelle, M., Booktein, F. L., Braga, J., Halazonetis, D. J., Weber, G. W. y Mitteroecker, P.
(2011) Sexual dimorphism of the human mandible and its association with dental development. American Journal of Physical Anthropology, 145, 192-202.
Dávalos Hurtado, E.
(1951) La Deformación Craneana entre los Tlatelolcas [Tesis de licenciatura en Antropología física y maestría en Ciencias antropológicas]. Instituto Nacional de Antropología e Historia/Universidad Nacional Autónoma de México.
Enlow, Donald H.
(1990) Facial Growth. W. B. Saunders Company, Philadelphia, Pennsylvania, USA.
García Moll, R.
(1977) Análisis de los materiales arqueológicos de la cueva del Texcal, Puebla. Colección Científica No. 56. Arqueología. Departamento de Prehistoria, Instituto Nacional de Antropología e Historia-Secretaría de Educación Pública: México.
González-José, R., Dahinten, S. L., Luis, M. A., Hernández, M. y Pucciarelli, H. M.
(2001) Craneometric variation and the settlement of the Americas: testing hypotheses by means of R-matrix and matrix correlation analyses. American Journal Physical of Anthropology, 116, 154-165.
González-José, R., Neves, W., Mirazon-Lahr, M., González, S., Pucciarelli, H., Hernández-Martínez, M. y Correal, G.
(2005) Late Pleistoce/Holocene craniofacial morphology in Mesoamerican Paleoindians: implications for the peopling of the New World. American Journal Physical of Anthropology, 128, 772-780.
González, S., Jiménez, J. C., Hedges, R., Huddart, D., Ohman, J.C., Turner, A. y Pompa, J. A.
(2003) Earliest human in the Americas: new evidence from Mexico. Journal of Human Evolution, 44, 370-387.
González, A., Rojas, C., Terrazas, A., Benavente, M. E., Stinnesbeck, W., Avilés, J., de los Ríos, M. y Acevez, E.
(2008) The arrival of humans on the Yucatan Peninsula: Evidence from
sumerged caves in the state of Quintana Roo, Mexico. Current Research in the Pleistocene, 25, 1-24.
Härdle, Wolfgang y Simar, Léopold
(2007) Applied Multivariate Statistical Analysis. Springer Berlin Heidelberg New York.
Hernández Flores, Rocío
(2018) Análisis de la morfología craneofacial en los primeros pobladores de México y su implicación en el poblamiento de América [Tesis de doctorado en Antropología, Instituto de Investigaciones Antropológicas, Facultad de Filosofía y Letras] Universidad Nacional Autónoma de México.
Hernández, R. y Martínez, G.
(2008) Morfología de la mandíbula de diferentes poblaciones de la cuenca de México (del Preclásico a la época Moderna) [Tesis de licenciatura en Antropología Física] Escuela Nacional de Antropología e Historia.
Hubbe, M., Harvati, K. y Neves, W.
(2011) Paleoamerican morphology in the context of European and East Asian Late Pleistocene variation: implications for human dispersion into the New World. American Journal of Physical Anthropology, 144, 442-453.
Kaifu Y., Aziz, F. y Baba, H.
(2005) Hominid mandibular remains from Sangiran: 1952-1986 collection. American Journal Physical Anthropology, 128, 497-519.
Jantz, R. L. y Douglas W. O.
(2001) Variation among early North American crania. American Journal of Physical Anthropology, 114, 146-155.
Jiménez, J. C. y Hernández, R.
(2011). Mujer de Texcal. En: J.C. Jiménez, C. Serrano, A. González y Felisa J. Aguilar (Coord.) IV Simposio Internacional El Hombre Temprano en América (pp. 107-120), Instituto de Investigaciones Antropológicas-Universidad Nacional Autónoma de México-Instituto Nacional de Antropología e Historia-Museo del Desierto A.C.: México.
Jiménez, J. C., Hernández R., Martínez G. y Saucedo, G.
(2006) La mujer del Peñón III. En: J. C. Jiménez, S. González, J. A. Pompa y F. Ortiz (Coords.), El Temprano en América y sus implicaciones en el poblamiento de la cuenca de México. Primer Simposio Internacional (pp. 49-66). Colección Científica. Serie Antropología Física. Instituto Nacional de Antropología e Historia.
Jiménez, J. C., Hernández, R. y Martínez, G.
(2009) La morfología de los primeros humanos de la cuenca de México. Ciencia, Conocimiento y Tecnología. Un Polizón Llamado Darwin, 90, 68-71.
Jiménez, J. C., Martínez, G. y Hernández, R.
(2014) Primeras evidencias humanas en la cuenca de México. En Eduardo Corona M. y Joaquín Arroyo-Cabrales (Coords.), Perspectivas de los estudio de Prehistoria en México. Un Homenaje a la trayectoria de Joaquín García Bárcena. Colección Arqueología, Serie Logos, Instituto Nacional de Antropología e Historia.
Jiménez, J. C., Pompa, J. A., Hernández, R. y Martínez, G.
(2003) Los primeros pobladores de Santa María Aztahuacan. Cincuentenario. UNAM, FES-Zaragoza, ENTS, CONACULTA, INAH, Delegación Iztapalapa y el Voluntariado Social de Iztapalapa.
Jiménez, J. L., Tlalolini, N. E., Salas, E. G., Romero, S. E., Zárate, P. y Fong, R.
(2016) Nuevos fechamientos en la cuenca de México. Ponencia presentada en el VIII Simposio Internacional El Hombre Temprano en América, Museo de Antropología de Xalapa.
Johnson, F. y R. S. MacNeish
(1972). Chronometic Dating. En D. S. Byers (Ed.), The Prehistory of the Tehuacan Valley, vol. 4 (pp. 3-58), Austin.
Lagunas Rodríguez, Zaid
(1967) Estudio métrico y morfológico de mandíbulas prehispánicas de México (Tlatelolco) [Tesis de licenciatura, Escuela Nacional de Antropología e Historia, SEP/INAH].
Lieberman, Daniel E.
(2008). Speculations about the selective basis for modern human craniofacial form. Evolutionary Anthropology, 17, 55-68.
Lieberman, D. E.
(2011) The Evolution of the Human Head. The Belknap Press of Harvard University Press.
Lieberman D. E., Brandeis, M. M. y Krovitz, G.
(2002) The evolution and development of cranial form in Homo sapiens. PNAS, 99, 1134-1139.
Lovejoy C. O., Meindl, R. S., Pryzbeck, T. R. y Mensforth, R. P.
(1985) Chronological metamorphosis of the auricular surface of the ilium: A new method for the determination of adult skeletal age at death. American Journal Physical of Anthropology. 68, 15-28.
Loth, S. R. y Henneberg, M.
(1996) Mandibular ramus flexure: a new morphologic indicator of sexual dimorphism in the human skeleton. American Journal Physical Anthropology, 99, 473-485.
Martínez-Abadías N., Esparza, M., Sjovold, T., González-José, R., Santos, M., Hernández, M. y Klingenberg, C. P.
(2011) Pervasive genetic integration directs the evolution of human skull shape. Evolution, 64(4), 1010-1023.
Meindl, R. S. y Lovejoy, C. O.
(1985) Ectocranial suture closure: A revised method for the determination of skeletal age at death based on the lateral-anterior sutures. American Journal Physical of Anthropology, 68, 57-66.
Mirazon-Lahr, M.
(1995) Patterns of modern human diversification: Implications for Amerindian origins. American Journal Physical Anthropology, 38, 163-198.
Morant, G. M.
(1936) A biometric study of the human mandible. Biometrika, 28, 84-122.
Morett, L., López, D. y Ramírez, B.
(2004) Acercamiento al hombre de Texcoco a través del análisis osteológico. Nuestro Espacio Universitario, 9, 25-26.
Neves, W. A. y Pucciarelli, H.
(1989) Extra-continental biological relationships of early South American human remains: a multivariate analysis. Ciência e Cultura, 41, 566-575.
(1991). Morphological affinities of the first Americans: and exploratory analysis based on early South American humans remains. Journal of Human Evolution, 21, 261-273.
Neves, W. A., Powell, J. F. y Ozolins, E. G.
(1999) Extracontinental mophological affinities of Palli Aike, Southern Chile. Interciencia, 24, 258-263.
Neves, W.A., González-José, R., Hubbe, M., Kipnis, R., Araujo, A. G. M. y Blasi, O.
(2004) Early Holoceno human skeletal remains from Cerca Grande, Lagoa Santa, Central Brazil, and the origins of the first Americans. World Archaeology, 36, 479-501.
Neves, W. A. y Martínez, M. M.
(2005) Afinidades biológicas de grupos pré-históricos do vale do rio Ribeira de Iguape (SP): uma análize preliminar. Revista de Antropología, 48, 525-558.
Pompa y Padilla, José A. y Serrano Carreto, E.
(2001) Los más antiguos americanos. Arqueología Mexicana. IX(52), 36-41.
Powell, J. F. y Neves, W. A.
(1999) Craniofacial morphology of the first Americans: pattern and process in the peopling of the New World. Yearbook of Physical Anthropology, 42, 153-188.
Powell, J. F., Neves, W. A., Ozolins, E. y Pucciarelli, H. M.
(1999) Afinidades biológicas extracontinentales de los dos esqueletos más antiguos de América: implicaciones para el poblamiento del Nuevo Mundo. Antropología Física Latinoamericana, 2, 7-22.
Pucciarelli, H. M.
(2004) Migraciones y variación craneofacial humana en América. Complutum, 15, 225-247.
(2009) Explicando el paleoamericano y su probable extinción, a través de inferencias comparativas y experimentales. RUNA XXX, 1, 9-27.
Pucciarelli, H. M., Sardi, M. L. y Jiménez, J. C.
(2003) Poblamiento temprano de América y diversificación evolutiva. Estudios de Antropología Biológica, XI, 661-678.
Püschel, T.
(2014) Modularidad e integración morfológica en cráneos humanos: un enfoque morfométrico geométrico. International Journal of Morphology, 32, 299-304.
Rodríguez, J. V. (Ed.)
(2001) Craneometría de la población prehispánica de los Andes Orientales de Colombia: diversidad, adaptación y etnogénesis. Implicaciones para el poblamiento americano. Los Chibchas, (pp. 51-310.). Universidad Nacional.
Romano, A.
(1955) Nota preliminar sobre los restos humanos sub-fósiles de Santa María Astahuacán. Anales del INAH, 36, 65-74.
(1963). Breve informe de los hallazgos en San Vicente Chicoloapan, México. Sobretiro de los Anales del INAH, XV, 245-259.
(1970) Preceramic human remains. En Stewart T. D. (Ed.), Handbook of Middle American Indians (pp. 22-34). University of Texas Press Austin.
(1974) Restos óseos precerámicos de México. En: J. Romero (Coord.), Antropología física, Época Prehispánica. México: panorama histórico y cultural (pp. 29-81). Instituto Nacional de Antropología e Historia.
Rosas, A.
(2001) Occurrence of neandertal features in mandibles from the Atapuerca – SH site. American Journal of Physical Anthropology, 114, 74-91.
Rosas, A. and Aguirre, E.
(1999) Restos humanos neandertales de la cueva del Sidrón, Piloña, Asturias. Estudios Geológicos, 55, 181-190.
Salas, M. E., Pijoan, C. M. y García, R.
(1988) Estudio comparativo de los restos fósiles humanos localizados en México. En J. Alba González (Comp.), Orígenes del Hombre Americano (pp. 127-144). Secretaría de Educación Pública.
Serrano, C., Hernández, R. y Gómez-Valdés, J. A.
(2016) Nuevo dato radiocarbónico de un esqueleto del Holoceno temprano procedente de Tláhuac, Ciudad de México. Ponencia presentada en el VIII Simposio Internacional El Hombre Temprano en América, Museo de Antropología de Xalapa, Veracruz.
Terrazas, Alejandro y Benavente, Martha E.
(2006) Estudio preliminar de tres cráneos tempranos, procedentes de cuevas sumergidas de la costa este de Quintana Roo. En J. C. Jiménez, O. J. Polaco, G. Martínez y R. Hernández (Coords.), Segundo Simposio Internacional El Hombre Temprano en América (pp. 189-197). CONACULTA-INAH.
Ubelaker, D. H.
(1978). Human Skeletal Remains: excavation, analysis, interpretation. Washington, Taraxacum.
Walker, M. J., Lombardi, A. V., Zapata, J. and Trinkaus, E.
(2010) Neandertal mandibles from the Sima de las Palomas del Cabezo Gordo, Murcia, Southeastern Sapain. American Journal of Physical Anthropology, 142,
261-272.
White T. D., G. Suwa y B. Simpson
(2000) Jaws and teeth of Australopithecus afarensis from Maka, Middie Awash, Ethiopia. American Journal of Physical Anthropology, 111, 45-68.
White, T. D. y Folkens, P. A.
(2005) The Human Bone Manual. El Servier Academic Press.
1 Se debe mencionar que algunos de ellos sólo cuentan con dataciones indirectas o estimadas.
2 Una cabeza dolicomórfica determina una cara estrecha, larga y protrusiva, con un paladar de modo correspondiente, más largo, estrecho y profundo, y a la inversa, una cabeza braquimórfica establece una cara amplia y menos protrusiva, que determina un paladar y arco dental más ancho y corto. En este sentido, el arco mandibular está relacionado con la arcada superior, o dicho de otra forma, el cuerpo de la mandíbula es la contraparte estructural del cuerpo del maxilar, y sus longitudes, así como su posición, se encuentran en equilibrio (Enlow, 1990).
3 Solo para aquellos casos en donde se contó con las medidas requeridas para los cálculos de estos índices.
4 La base del cráneo está conformada por el occipital, temporal, esfenoides y etmoides.
